ВIЗЕРУНЧАСТИЙ ПОЛОЗ
Є.В.Зав▓ялов
Dione snake in Povolzhye
The article reports on today▓s distribution of the dione snake. The information comes from literature, study of the museum collections and modern field researches, carried out by the authors in the Lower Volga area in 1998-2000. The data on species biology include the population number and density, reproduction (mating, development and incubation of eggs), so as food preferences.
Вiзерунчастий полоз √ один iз найбiльш поширених видiв самоп чисельноп родини змiй
Colubridae. Незважаючи на широкий євроазiатський ареал та велику iсторiю вивчення вiзерунчастого полоза, внутрiшньовидова диференцiацiя виду недостатньо розроблена, а вирiшення питань пiдвидовоп дiагностики носить i нинi дискусiйний характер. Пiдтвердженням цьому служить чимало синонiмiчних назв географiчних форм даних полозiв, описаних рiзними авторами до теперiшнього часу. Так, полози iз окремих районiв (Захiдного Сибiру та Алтаю) цього широкого ареалу неодноразово описували в ранзi окремих пiдвидiв (Elaphe dione niger Golubjeva, 1923) та (E.dione tenebrosa Sobolevsky, 1929), з характерним дуже темним, без усякого рисунку, забарвленням тiлом. Подальшi дослiдження показали, що цi форми не мають таксономiчного значення i зведенi в синонiми E.dione (Терентьев, Чернов, 1949). Окрiм того, описаний О.М.Никольським (1914) iз Примор΄я Coluber cherskii був також визнаний варiєтетом E.dione (Павлов, 1926). Однак тепер ряд систематикiв (Obst, Scerbak, 1993) знову визнають можливим видiлити пiдвидову самостiйнiсть популяцiй вiзерунчастого полоза iз Далекого Сходу (Примор΄я). Як було показано ранiше (Емельянов, 1929; Strauch, 1873), цi полози вiдрiзняються не тiльки особливостями рисунку та забарвленням, но i харатерезуються меншим числом черевних та пiдхвостових щиткiв; при отриманнi достовiрних даних, пiдтверджуючих це припущення, далекосхiдний пiдвид, явно, повинен мати назву E.d.cherskii (Obst, Scerbak, 1993).İñ
нує й iнша точка зору, згiдно якої варiабельнiсть морфометричних ознак у вибiрках полоза iз Далекого Сходу вписується в межi мiнливостi виду iз цього великого ареалу.Ареал виду простягається вiд Кореї, Примор΄я, Пiвнiчного Китаю та Далекого Сходу через Центральну i Середню Азiю до Лiвобережя України, Пiвнiчного İрану та Закавказ΄я (Ананьева и др., 1998; Obst, Scerbak, 1993; Zhao, Adler, 1993; Brunner, 1995; Schulz, 1996; Darevsky, 1997). На пiвденному заходi Россiї вiзерунчастий полоз зустрiчається скрiзь; тут пiвнiчна границя розповсюдження проходить по територiях Воронежської, Ульяновської, Самарської, Саратовської та Оренбургської областей (Гаранин, 1983; Бакиев, 1998; Табачишин, Завьялов, 2000).
При дослiдженнi полозiв у Поволжi встановлено, що найбiльша довжина тулуба з хвостом досягає 1031.0 мм. У самцiв хвiст у 3.83-4.35, а у самок - 4.25-5.87 раза менший тулуба. Голова звужена до морди, найбiльшої ширини досягає бiля кутiв рота. Мiжщелепний щиток тупим кутом вдається помiж мiжносовими. Нiздрi округленi, розташованi мiж двома щитками. Передорбiтальний щиток 1, пiдорбiтальних - 1 (23.0%) або 2 (77.0%), позаорбiтальних - 2 (93.9%) та 3 (6.1%), вискових √ 2+3 (71.7%), 2+4 (14.
3%), 3+3 (6.6%), 3+4 (3.8%) та 2+2, 2+5, 3+2, 3+5 по 0.9%. Верхньогубних щиткiв 8/8 (88.9%), або 9/8 + 8/9 (9.3%) та 9/9 (1.8%), iз яких 4-й або 5-й торкаються ока. Спиннi лусочки посерединi спини з невиличкими, але добре помiтними реберцями, на боках тулуба вони гладенькi. Анальний щиток роздiлений.Забарвлення верхньої сторони тiла буро-сiре (рис. 1). На головi видiляється характерний для виду рисунок, який складається iз поперечної, дугоподiбної темної з чорними краями смуги. Вона проходить по заднiх частинах перед-чiльному та передньої частини чiльного щиткiв i закiнчується бiля передньоверхнього краю ока. По боках голови вiд заднього краю ока до кутiв рота iде бура, вiдмежована чорним смуга. У статевозрiлих полозiв спинна сторона, як правило, сiруватого або сiровато-бурого кольору, з чотирма повздожними бурими або буруватими смугами вздовж тулуба. На хребтi вузькi, неправильнi, поперечнi, темно-бурi або чорнi смуги, промiжки мiж якими бiльшi їх ширини у 4-6 разiв. Черево сiрувате або блiдочервоновате з чорними яркими або бурими, неправильно, щiльно розташованими плямами. Забарвлення молодих полозiв як у дорослих, але значно яскравiше.

Рис. 1.
Вiзерунчастий полоз один iз найбiльшСамки бiльшi самцiв, а вiдносна довжина голови у них менша по порiвнянню з самцями. У самок, крiм цього, бiльше черевних щиткiв, но менше пiдхвостових щиткiв (p
< 0.05). Таким чином, половий диморфiзм виявляється по довжинi тулуба, iндексу хвоста, числу черевних та пiдхвостових щиткiв, вiдношенню довжини голови до довжини пiлеусу та вiдношенню найбiльшої довжини чiльного щитку до вiдстанi вiд його переднього краю до до шову мiж мiжносовими та мiжщелепними щитками.Зiставлення даних по фолiдозу плазунiв iз Поволжя та даних лiтератури iз iнших регiонiв в межах ареалу, значних у пiдвидовiй систематицi вiдминностей не виявлено.
Вiзерунчастий полоз вiдноситься до еврiбiонтних видiв з високою пластичнiстю при виборi бiотопу. Звичайними бiотопами виду на данiй територiї являються узлiсся нагiрних дiбров, чагарниковi заростi, обриви по берегах водоймищ та схили понижень ярково-балкових систем (рис. 2). Дуже часто вони спостерiгаються i в антропогенному ландшафтi: фруктових садах, зрошуваних землях та рисових полях, руїнах старих будiвель, населенних пунктах.
Численнiсть вiзерунчастого полоза в умовах Самарської областi (в де яких бiотопах Нацiонального парку ⌠Самарська лука■) становила у минулому 7.0 i навiть 11.0 ос./км маршруту (Баринов, 1982). Нинi на цiй територiї найбiльша густота виду не перевищує 3.0 ос./га (Бакиев, 1998). У Лiвобережi Саратовської, Волгоградської та Астраханської областей змiя зустрiчається скрiзь; великi показники густоти виду виявленi у долинах рiк Волги, Великого i Малого Узенiв та їх притокiв (Табачишин, Завьялов, 2000). Густота населення виду на пiвночi областi поширення коливається вiд 0.8√1.2 ос./га (на дiлянках рiзнотравно-типчакового степу та його похiдних у Балаковському i Пугачовському районах Саратовської областi) до 28.4 ос./га (на окремих дiлянках о.Круглий у середнiй зонi Волгоградського водосховища у Ровенському районi). На пiвднi європейської частини РФ в Астраханськiй областi щiльнiсть вiзерунчастого полоза коливається вiд 0.62 та 1.28 ос./га (на дiлянках
горбистих ланцюгових пiскiв, закрiплених полинно-злаковою рослиннiстю з солончаками у пониженнях) до 18.2 ос./га (на закрiплених горбистих пiсках).
Рис. 2.
Нори ластівки берегової - улюблене мiсцеУ Правобережi на значних дiлянках територiї вiзерунчастий полоз являється звичайним, однак на пiвночi регiону рiдкiсний. Наприклад, в травнi 1996 р. в межах Воскресеньського району Саратовської областi на узлiссi заплавнего лiсу гирла р.Чардим спостерiгалась 1-2 особини на 1 км маршрутного дослiдження, на схилi рiчкової тераси р.Волга бiля с.Усовки Саратовського району
- 2.5. По даним абсолютного облiку (05.08.1997р.) на окремiй дiлянцi в степу (2110 м2) бiля центральної садиби Балабанiвського мисливського господарства (Чернишковський р-н Волгоградської областi) вiдмiчено 5 полозiв, що становить 23.7 ос./га.В умовах Поволжя активнi полози спостерiгаються з першої половини березня по другу половину жовтня, iнколи до середини листопада. Перша поява їх зареєстрована у гирлi р.Уси (Самарська обл.) 14.04.1995р. (Бакиев, 1998), на околицях м.Саратова 18.03.1995 р., бiля озера Ельтон (Волгоградська обл.) 15.03.1949 р. (Косарева, 1950).
Вiзерунчастий полоз активний вдень незалежно вiд пори року. Лiтом у цього полоза цикл активностi двовершинний, коли видiляються ранковий та вечiрнiй пiки, тодi як весною та восени вiн одновершинний. Так, з першої половини червня активнi полози на пiвднi Саратовської та Волгоградської областей, як правило, спостерiгались з 8 до 12 г ранку та 17 до 20 г вечером. Анологiчнi закономiрностi виявленi для вiзерунчастого полоза у Казахстанi (Параскив, 1956), Монголiї (Ананьева и др., 1997) та Українi (Таращук, 1959). Температура на поверхнi субстрата у перiод найбiльшої активностi змiй коливається вiд 22.4 до 33.1╟С, а ректальна температура тiла √ 19.8-34.7╟С.
Шлюбний перiод у тварин починається незабаром пiсля пробудження. Вiдкладання яєць вiдбувається наприкiнцi червня або у серединi липня; такi кладки були виявленi С.Черновим 23.06.1950 р. у Волго-Уральському межирiччi (Чернов, 1954) та 15.07.1997 р. i 10.07.1998 р. бiля с.Ровно Саратовської областi. Мiж iншим, в умовах терарiума спостерiгалось парування полозiв i восени. Таке явище, повязано з тривалiстю перiоду збереженя житнєздатностi сперматозоїдiв, характерної для популяцiй полозiв, що живуть у несприятливих клiматичних умовах (Schulz, 1995).
Тривалість розвитку та iнкубацiї яєць залежать вiд екологiчних умов бiотопу тварин та клiматичних умов перiоду. Наприклад, у двох самок, вiдловлених 30.06.1998 р., виявлено по 8 яєць, при розкриваннi яких виявлено цiлком сформованi, але без лускового покрову зародки довжиною 14 мм. Кiлькiсть яєць у кладцi коливається вiд 8 до 14. Їх розмiри (
n= 40) 18.2-25.0ґ 36.0-46.1 мм (у середньому 20.8╠ 0.21ґ 40.7╠ 0.32). Молодi полози вилуплюються з яєць при температурi 23.5-31.9╟С через 23-28 (у середньому 25.1) дiб.
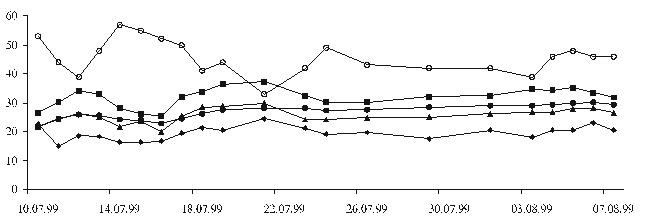
Рис. 3.
Динамiка экологiчних факторiв у перiод iнкубацiї яєць вiзерунчастого полозаДовжина тулуба та хвоста щойно народжених полозiв досягає (
n=12) 211.0-230.0 та 43.0-54.1мм вiдповiдно (у середньому вiдповiдно 217.1╠ 1.28 та 47.8╠ 0.67 мм). Пiсля першого линяння приблизно на 4-6 добу молодi тварини починають активне полювання.Згiдно з лiтературними даними головну поживу вiзерунчастого полоза становлять дрiбнi гризуни, птахи та їх яйця, плазуни. В цiлому для виду на дослiджуванiй територiї характерний широкий спектр поживних об΄єктiв. Так, на пiвднi Саратовської областi (околиця с.Ровно) та у межирiччi Великого та Малого Узенiв (Олександрово-Гайський район) у шлунках змiй були виявленi рештки гризунiв (
Microtus sp.), прудкої ящiрки (Lacerta agilis), пташок та яєць ластiвки берегової (Riparia riparia). По даним В.Шапошнiкова та В.Жукова (1988), у Самарськiй областi полоз живиться гризунами (87.5%), ящiрками (8.4%), яйцями та горобцеподiбними птахами (4.1%); анологiчнi данi вiдомi для Волгоградської областi (Косарева, 1950) та Волго-Уральського межирiччя (Чернов, 1954).БIБЛIОГРАФIЧНИЙ СПИСОК
Ананьева Н.Б., Боркин Л.Я., Даревский И.С., Орлов Н.Л. Земноводные и пресмыкающиеся. Энциклопедия природы России. М., 1998. С. 1-576.
Ананьева Н.Б., Мунхбаяр Х., Орлов Н.Л., Тэрбиш Х. Семейство Ужеобразные √
Colubridae Oppel, 1811 // Земноводные и пресмыкающиеся Монголии. М., 1997. С. 281-325.Бакиев А.Г. Эколого-фаунистические исследования змей Среднего Поволжья, экологические основы охраны офидиофауны и рационального использования ядовитых видов в регионне. Автореф. дис. ...канд. биол. наук. Н.Новгород, 1998. 23 с.
Гаранин В.И. Земноводные и пресмыкающиеся Волжско-Камского края. М., 1983. 176 с.
Емельянов А.А. Змеи Дальнего Востока // Зап. Владивосток. отд. рус. геогр. об-ва. Владивосток, 1929. Т. 3 (20), вып. 1. С. 1-208.
Косарева Н.А. Рептилии юга Сталинградской области // Учен. зап. Сталинград. пед. ин-та. Cталинград, 1950. С. 227-240.
Никольский А.М. Новые для русской фауны пресмыкающиеся из южной части Приморской области // Ежегодн. Зоол. Муз. Имп. Акад. наук. 1914. Т. 19, ╧1. С. 88-91.
Никольский А.М. Фауна России и сопредельных стран. Пресмыкающиеся (
Reptilia). Пг., 1916. Ч. 2. 349 с.Павлов Б.А. Животный мир Маньчжурии по коллекциям Общества изучения Маньчжурского края (Пресмыкающиеся и земноводные) // Вестник Маньчжурии (Секция естествознания. Сер.А. Вып. 13). Харбин, 1926. ╧8. С. 1-22.
Параскив К.П. Пресмыкающиеся Казахстана. Алма-Ата, 1956. 227 с.
Таращук В.I. Плазуни // Фауна України. Київ, 1959. Т. 7. 246 с.
Терентьев П.В., Чернов С.А. Определитель пресмыкающихся и земноводных. М.-Л., 1949. 340 с.
Чернов С.А. Эколого-фаунистический обзор пресмыкающихся юга междуречья Волга-Урал // Труды Зоол. ин-та АН СССР. 1954. С. 137-158.
Шапошников В.М., Жуков В.П. Охрана узорчатого полоза на Самарской Луке // Охрана животных в Среднем Поволжье. Куйбышев, 1988. С. 25
-29.Brunner M. Morphologische Analyse der palaarktischen Schlangengruppe Elaphe dione (Reptilia, Serpentes). Diplomarbeit. Zoolog. Museum der Univ. Zurich, 1995. 88 p.
Darevsky I.S. Elaphe dione (Pallas, 1773) // Atlas of Amphibians and Reptiles in Europe. Paris, 1997. P. 352, 353.
Obst F.J., Scerbak N.N. Elaphe dione (Pallas, 1773). Steppennatter // Handbuch der Reptilien und Amphibien Europas. AULA √ Verlag. Wiesbaden, 1993. P. 295-315.
Schulz K.D. A monograph of the colubrid snakes of the genus Elaphe Fitzinger. Koeltz Scientific Books. 1996. 439 p.
Strauch A.A. Die Schlangen des Russischen Reichs, in systematischer und zoogeographischer Beziehung geschildert // Mem. Acad. Imper. Sci. St.-Petersb., 1873. Ser. 7. Vol. 21, ╧4 (2). P. 1-287.
Zhao E.M., Adler K. Herpetology of China. Contribution to Herpetology. Soc. Study Amphib. Reptiles. 1993. ╧10. 522 p.
╘
Ukrainian magazine "Naturalist"