ЗМІНИ РИБОВОДНО-БІОЛОГІЧНИХ ПОКАЗНИКІВ
САМОК ТОВСТОЛОБИКІВ ЗАЛЕЖНО ВІД
КРАТНОСТІ ЦИКЛІВ ВІДТВОРЕННЯ
Опубліковано ISSN 0557-5672 "Рибне господарство" 1993 вип. ╧ 47, С.32-37;
Микола Рудь
Костянтин Рибальський
У роботі розглянуто актуальні питання поліциклічного розмноження
білого та строкатого товстолобиків, вирощених у садках у теплих водах.
Піддано аналізу пов'язані з цим зміни рибоводно-біологічних показників
самок обох идів товстолобиків.
Суттєвим фактором, який стримує поліпшення якості і масштабів виробництва личинок товстолобиків від плідників, вирощених у садках у теплих водах, є одноразове використання їх для відтворення з рибоводною метою (Балтаджі Р.А., 1984). Це зумовлено значним відходом вперше дозрілих плідників у післянерестовий період. Пошук шляхів підвищення кількості особин. Що вижили після відбору статевих продуктів, почали із широкого впровадження товстолобиків як об'єктів рибництва (Виноградов К.В., Єрохіна Л.В., 1974). Причинами загибелі товстолобиків після відбору статевих продуктів, очевидно, є травми лускового покриву, одержані на всіх етапах технологічного циклу відтворення, а також невідповідність стану понад дозуванню гормональних препаратів та часу взяття у риб статевих продуктів. Результатом порушення гормонального балансу є різка зміна фізіологічного стану організму риб, що проявляється в помутнінні очей, порушенні координації рухів, запаленні покривних тканин голови та, у кінцевому результаті, загибелі.
Розроблена нами методика прижиттєвого, нетравматогенного визначення готовності риб до нересту дає можливість у польових умовах за допомогою простого пристрою точно визначити стан понад, коректувати кратність та дозування гормональної ін'єкції (Рудь М.П., 1989). Це значною мірою сприяв зберіганню плідників, оберігає організм проін'єктованих особин від передозування, забезпечує у післянерестовий період добрий функціональний стан організму, скорочує строк наступного дозрівання нової генерації яйцеклітин. Крім цього, суттєво сприяє зберіганню та зумовлює швидке відновлення репродуктивних функцій організму використання біологічно активних та ранозаживляючих речовин.
Зберігання плідників дає можливість не тільки вирішувати проблеми багаторазового їх використання у рибоводних цілях, без якої селекція як найважливіший елемент культивування взагалі малоефективна, але й розпочати створення породної групи, стійкої до несприятливих умов промислового розведення, з прискореним дозріванням та нерестом у більш ранні строки (Кормилін В.В., Цой Р.М., 1982). Крім того, з'являється можливість зменшити або навіть виключити з числа цьогорічок, що залишають на плем▓я, особин, одержаних від плідників, що дозріли вперше.
В результаті досліджень 1986-1990 рр., крім збереження плідників та використання їх знову для відтворення, ставили за мету виявити особливості розвитку нової генерації яйцеклітин у післянерестовий період в умовах утримання в теплиї водах. По аналогії з роботами А.М.Багрова та В.Г.Чертихіна (1985), проведеними в умовах ставкових господарств о.Куба з білим товстолобиком , вважали за можливе багаторазове одержання потомства в одному вегетаційному від плідників білого та строкатого товстолобиків (БТ і СТ), вирощених у садках в умовах водойми-охолоджувача. Аналіз гістологічних проб, що періодично відбирались у плідників, що брали участь у нерестовій кампанії, показав можливість багаторазового отримання потомства. При цьому самці обох видів є текучими з квітня по жовтень при констатації задовільного стану сперми (колір, консистенція, рухомість сперміїв).
Такий подовжений період текучості самців в умовах теплих вод, вірогідно, зумовлений асинхронним характером сперматогенезу, приматанним рослиноїдним рибам (Горбач Є.І., 1965; Єрохіна Л.В., Виноградов В.К., Вичегжаніна В.В., 1976; Виноградов В.К., Єрохіна Л.В., 1977; Рибаков В.Ю., Балтаджі Р.А., Руденко А.П. та ін. 1987).
Методика досліджень. У наших дослідах по багаторазовому одержанню потомства від одних і тих
же плідників протягом одного вегетаційного сезону використовували 4-х річних самок (3+) середньою
живою масою БТ - 2260, СТ - 2360г, які були вирощені у водоимі-охолоджувачі Бурштинської ДРЕС
у садках розміром 3 X 4 м та об▓ємом 24 м3 з температурою води у весняно-літній період 26-34╟С,
а взимку не нижче 7╟С. Відбір статевих продуктів, інкубація ікри, витримування, підрощування та вирощування молоді проводили в рамках технологічних вимог. Матеріали, наведені в даній статті,
були піддані статистичній обробці.
Результати досліджень. Кількість розчиненого у воді кисню в районі садкової лінії становила в середньому 5,5 мг/л, не знижуючись протягом року нижче 4,0 мг/л. У результаті досліджень розвитку кормової бази водойми-охолоджувача, проведених попередніми роками, встановлено, що з травня по жовтень біомаса фітопланктону знаходилась у межах від 3,14 до 5,8 г/м3. У травні в середньому по водоймі вона становила 1,7 г/м3. Близькі показники за цей період відмічені у районі скидного каналу в місці розташування садкової лінії. Влітку (серпень) середня маса зоопланктону зменшувалася до 0,7 г/м3 і у жовтні становила 0,2 г/м3. Така динаміка розвитку кормової бази з травня по жовтень характерна для даної водойми і має тенденцію до щорічного повторення з незначними відхиленнями (Носаль О.Д., 1972; Носаль О.Д., Тарасова О.М., Балтаджі Р.А. та ін., 1974; Данченко А.Д., Сидоров Н.А., 1988).
Із загальної кількості самок, яких використовували з метою відтворення у травні 1990 р., було збережено і використано в наступних дослідах 22 екз. строкатого та 12 екз.. білого товстолобиків, від яких 12.07.90 р. знову одержали ікру. І далі від зазначених самок строкатого товстолобика втретє 27.08.90 р одержали потомство. Таким чином, від самок білого товстолобика ікру одержали двічі, а від самок строкатого товстолобика - тричі в одному вегетаційному сезоні (з травня по серпень). При цьому самки із загальної кількості відібраних після першого розмноження і одержання ікри з метою збереження були знову двічі використані для розмноження і висаджені для наступного утримання в окремий садок. До 10 10.90 р. молодь білого товстолобика першої генерації характеризувалася масою 120, строкатого товстолобика - 115 г; молодь білого товстолобика другої генерації - 3,45, строкатого тоьстолобика - 2,97 г; молодь строкатого товстолобика третьої генерації мала масу 157,57 мг. Період зимівлі проходив при повній відсутності гедівлі. Відхід молоді при цьому не перебільшував 10 %. Друга генерація білого товстолобика на 08.05.91 р. характеризувалася середньою масою 15, строкатого - 14,3 г; третя генерація строкатого товстолобика - 3 г. Таке збільшення маси молоді за період зимівлі можна пояснити міграцією різних форм зоопланктону у зону, що прилягає до місця скидання теплих вод електростанції, де розташована садкова лінія.
Високий рівень розчиненого у воді кисню і відносно висока температура води, можливо, є причиною міграції й концентрування у цій ділянці водойми кормових організмів і, як наслідок, достатньо високого темпу росту молоді у зимовий період.
Багаторазове використання садкових плідників для відтворення в одному сезоні дало можливість визначити динаміку змін рибоводно-біологічних показників (рис. 1-8). При цьому між одержанням першої
і другої генерації сума активних температур становила 1600 гр/дн, а між другою і третьою - 1200 гр/дн.
.JPG)
1. Динаміка зміни показника робочої
плодючості:
І - перша генерація потомства від плідників,
котрі нерестували вперше, одержана 14.05.90 р.;
ІІ - друга генерація потомства
одержана 10-13.07.90 р.;
ІІІ - третя генерація потомства
одержана 2.09.90 р.;
.JPG)
2. Динаміка зміни показника
відносної робочої плодючості:
1 - білий товстолобик;
2 - строкатий товстолобик;
.JPG)
3. Динаміка зміни виходу вільних
ембріонів з ікри:
1 - білий товстолобик;
2 - строкатий товстолобик;
.JPG)
4. Динаміка зміни маси одержаної ікри:
1 - білий товстолобик;
2 - строкатий товстолобик;
.JPG)
5. Динаміка зміни кількості ікринок в 1 г :
1 - білий товстолобик;
2 - строкатий товстолобик;
.JPG)
6. Динаміка зміни середньоарифметичного
показника діаметра овульованих ікринок:
1 - білий товстолобик;
2 - строкатий товстолобик;
.JPG)
7. Динаміка зміникоефіцієнта варіації
діаметру овульованих ікринок;
1 - білий товстолобик;
2- строкатий товстолобик;
.JPG)
8. Зміна показників виходу вільних ембріонів та
коефіцієнта варіації овульованих ікринок у самок,
розмножених одноразово у 1986 р. , і знову
використаних у 1987 р.:
1 - показник виходу вільних ембріонів;
2 - середньоарифметичний показник коефіцієнта
варіації діаметра овульованих ікринок
Зміна таких показників, як робоча плодючість (РП), відносна робоча плодючість (ВРП) *, процент виходу вільних ембріонів (% ВВЕ), кількість ікринок у 1 г, середнє арифметичне значення діаметру ікринок (Х О), маса відданої ікри, коефіцієнт варіації діаметру ікринок (Сv O) свідчить як про зміну умов утримання, так і про фізіологічні зміни, котрі відбулися в організмі самок після нерестової кампанії (див. рисунки 1-8).
Порівнюючи показники, слід відмітити, що білий та строкатий товстолобики мають деякі відмінності у рибоводно-біологічній характеристиці, які зумовлені різним спектром годівлі. Так, якщо графіки РП, ВРП, X О, кількості ікринок у 1 г, маси одержаної ікри у двох видів схожі, то % ВВЕ, залишаючись незмінним у білого товстолобика - 65,5-66%, різко змінюється у строкатого - 67,7-52-53,8 %. Найбільш суттєву різницю в обох видах спостерігали у Сv О, що зумовлено, очевидно, фізіологією процесів овогенезу й відмінністю у забезпеченні кормами в післянерестовий період. Спочатку більш високий Сv О у білого товстолобика змінюється у напрямі зменшення - 7,46 - 5,59, і навпаки - у строкатого цей показник різко збільшується від першого циклу розмноження до другого і потім набуває незначної зміни - 5,74 - 8,17 - 9,28. Пояснити це, напевно, можна незадовільними умовами годівлі, які, як було вже зазначено, з травня по жовтень за рівнем розвитку зоопланктону різко погіршуються.
По фітопланктону, який є основним кормом білого товстолобика, таких різких змін не відмічено.
Певно, це стало причиною того, що у фізіології процесу овогенезу негативних змін не спостерігали.
Слід зазначити, що Сv О є не стільки прямим, скільки відносним показником, котрий характеризує в більшій мірі варіативність форм ікринок, ніж діаметр ікри.
Можна вважати, що якість ікри товстолобиків перебуває у прямій залежності від вирівняності та розміру овульованих ікринок. Ікра однакової форми та розміру, як правило, має більш високі кондиції, що зумовлює високий процент запліднення, розвитку й виходу рухливих ембріонів. І навпаки: ікра різного розміру характеризується значною різноманітністю форми і, як правило, має невисокі кондиції (див. рисунки 3, 7).
Не маючи можливості показати, наскільки стабільна форма ікринок у тієї чи іншої особини, очевидно, можна користуватися показником варіативності діаметру, який характеризує,, таким чином, якість самки. Наприклад, у самок строкатого товстолобика, вирощених у 1987 р. у садках у водоймі-охолоджувачі Зміївської ДРЕС, яких використовували для розмноження у 1986 р. та збережених після цього, весною наступного року середній показник Сv О порівняно з попереднім мав значну тенденцію до зниження
- 7,14 - 5,01, що ілюструє взаємозв'язок коефіцієнту варіації діаметру ікринок з кратністю використання
в рибоводних цілях та з прогресом показника відсотка виходу вільних ембріонів відповідно 62 - 73%
(див. рисунок 8 ).
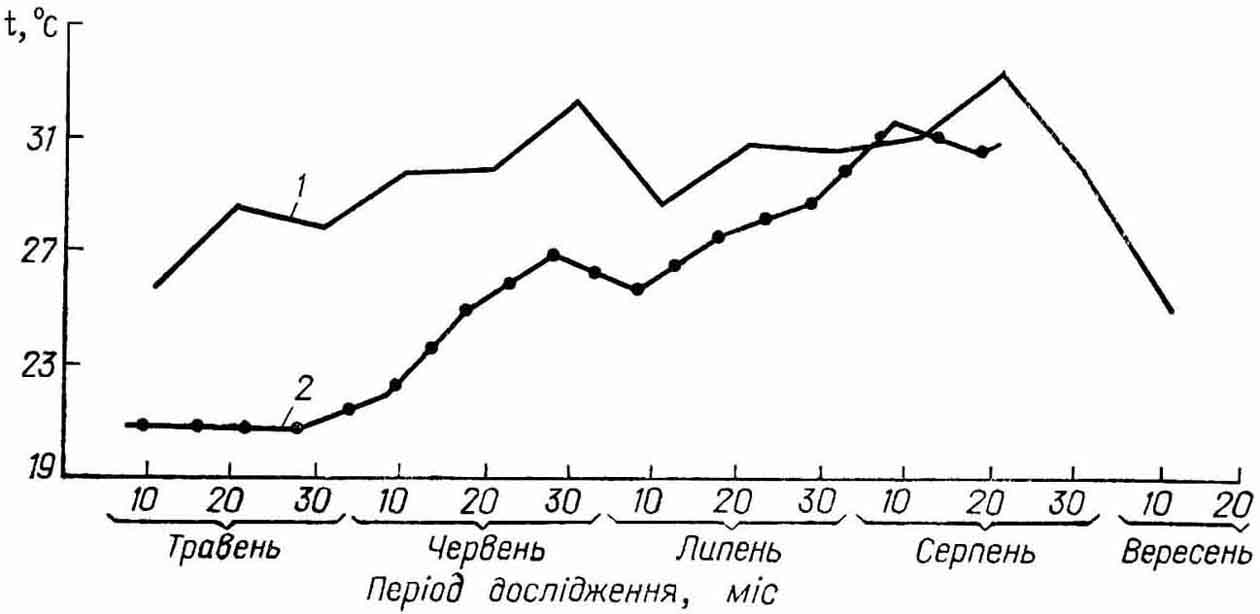
Загальна тенденція динаміки змін рибоводно-біологічних показників самок, яких використовували для поліциклічного відтворення, дає можливість припустити їх пряму залежність від рівня розвитку природної кормової бази. А також у випадку забезпечення плідників повним комплексом поживних речовин, визначає позитивну тенденцію зміни фізіологічних процесів та відтворення. Суттєвою особливістю овогенезу збережених самок є те, що весною наступного року вони дозрівають на два-три тижні раніше основного ядра маточного стада, яке складається з особин, що дозріли вперше. Таку ситуацію спостерігали в умовах водойми-охолоджувача Бурштинської ДРЕС, яка з кінця зими 1991 р. працювала в умовах зниженої потужності. Це зумовило різке зниження температури води у гирлі скидного каналу, що не
могло не вплинути на затримку готовності плідників до нересту (рис. 9).
Рис. 9 Динаміка температурного режиму водойми-охолоджувача
Бурштинської ДРЕС біля садкової лінії 1990 - 1991 рр.
графік 1 - 1990 р., графік 2 - 1991 р.
Тим більше, що збережені самки СТ, яких використовували тричі в попередньому 1990 р., дозріли на початок травня і 08.05.91 р. були відібрані для розмноження разом з буфало при середньонерестовій температурі води 20 ╟С.
У результаті гормонального стимулювання і від самців, і від самок нормально одержали статеві
продукти високої якості. У процесі інкубації температура води коливалася від 17╟С до 21╟С.
Проби ікри, що розвивалася, показали високий процент розвитку (95 %) рухомих ембріонів.
Виходячи з цього, важко прогнозувати кількість сезонів розмноження у поліциклічкому режимі без суттєвого зниження рибоводно-біологічних показників збережених плідників.
Проте, додаткові ресурси за рахунок поліциклічного відтворення товстолобиків, вирощених у садках
і експлуатованих в умовах тепловодного господарства, необхідно використовувати більш ефективно.
Висновки.
Збереження плідників після відбору статевих продуктів дає можливість використовувати їх два-три
рази протягом одного вегетаційного періоду.
Поліциклічне відтворення білого та строкатого товстолобиків можливе у господарствах, які базуються на водоймах-охолоджувачах енергетичних об'єктів при середньодобовій температурі з травня по серпень
25-З0╟С, із вмістом розчиненого у воді кисню не нижче 4,0 мг/л, розвитком фітопланктону від 3,14 до 5,8,
а зоопланктону - від 1,7 до 0,7 г/м3.
Аналіз змін динаміки рибсводно-біологічних показників дає можливість припускати її залежність
від рівня розвитку кормової бази.
Встановлено, що між першим і другим дозріваннями плідників сума активних температур становила
1500-1700, а між другим і третім - всього 1200 гр/дн.
Молодь другої і третьої генерації на початок травня наступного сезону досягає відповідно 14 і 3 г
і може бути використана у виробництві як маломірний посадковий матеріал.
Поліциклічне відтворення товстолобиків сприяє більш ранньому дозріванню збережених самок
весною наступного року, ніж особин, що вперше досягли статевої зрілості.
Впровадження у технологію елементів поліциклічнсто відтворення у тепловодних господарствах
дасть можливість суттєво збільшити їх виробничу потужність.
________________________
* Під ВРП у даному випадку слід розуміти відношення кількості одержаної ікри (РП)
до маси тіла самки у грамах.
БІБЛІОГРАФІЧНИЙ СПИСОК
1. Багров А.М., Чертихин В.Г. Особенности полового созревания и нереста самок белого толстолобика
в водоемах тропической зоны // Сб. науч. тр. ВНИИПРХ.- 1985.- Вып. 44. С. 90-92.
2. Биологическое обоснование вселення растительноядных рыб в водоем-охладитель Бурштынской
ГРЭС./ А.Д.Носаль, О.М.Тарасова, Р.А.Балтаджи, Л.Й.Сяра //Рыб. хоз-во.- 1974. - Вып.19. - С. 41 - 45.
3. Виноградов В.К., Ерохина Л.В. Опыт использования пенициллина как средства, снижающего
гибель производителей растительноядных рыб в нерестовой кампании.
4. Виноградов В. К., Ерохина Л. В. Руководство по биотехнике выращивания производителей
и эксплуатации маточных стад растительноядных рыб // М.: ВНИИПРХ, 1977.- 63 с.
5. Горбач 9. Й. Созревание й размножение белого амура Сtenopharingodon idella в среднем Амуре
// Вопр. ихтиологии. - 1965.- 5, вып. З (36). - С. 426 - 441.
6. Ерохина Л.В., Виноградов В.К., Вычегжанина В.В. Сравнительная характеристика сперматогенеза растительноядных риб // Тр. ВНИИПРХ.-1976. - Т. 25. - С. 34 - 40.
7. К вопросу о типах гаметогенезе у самок и самцов растительноядных рыб, выращенных в условиях водоемов-охладителей ГРЭС/Ф.Ю.Рыбаков, Р.А.Балтаджи, А.П.Руденко, Д.В.Бублик
// Рыб. хоз-во - 1987. - Вып. 41. - С. 23-28.
8. Кормилин В.В., Цой Р.М. Направления и методы селекции белого толстолобика в Казахстане
// Сб. науч. тр. ВНИИПРХ.-1982.-Т.33.- С. 104-110.
9. Рудь Н. П. Применение уретроскопа для определения половой зрелости рыб
// Рыб. хоз-во.-1980.- ╧ 4. - С. 49-51.
Одержано редколегією міжвідомчого тематичного
збірника "Рибне господарство" - 05.03.92.
В работе рассмотрены актуальные вопросы полициклического размножения
белого и пестрого толстолобиков, выращенных в садках на теплых водах.
Подвергнуты анализу связанные с этим изменения рыбоводно-биологических
показателей самок обеих видов.
╘
Ukrainian magazine "Naturalist"